A New Mode of T cell Suppression by LAG-3 Immune Checkpoint
What are checkpoint inhibitors?
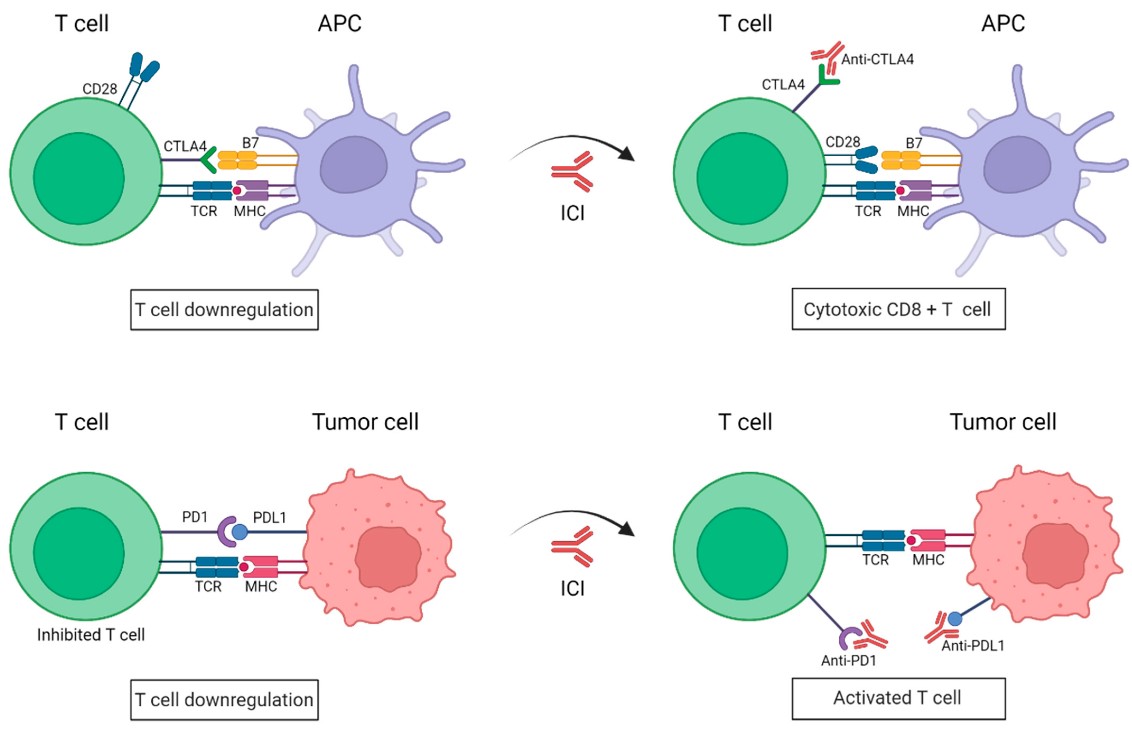
Checkpoint inhibitors, such as antibodies targeting the immune checkpoints programmed cell death-1 (PD-1) and cytotoxic T lymphocyte antigen-4 (CTLA-4) receptors expressed on T lymphocytes, are cancer immunotherapies that promote T-cell activation and tumor cell killing. Their efficacy is based on their capacity to prevent dendritic or tumor cells from interacting with T-cell receptors that would suppress activation and induce T-cell exhaustion.
Antibodies to PD1 and CTLA4 block immune checkpoints. Conventionally, binding of the T cell PD1 or CTLA4 receptors to their respective ligands (i.e., PDL1 on tumor cells or B7 on antigen-presenting cells, APCs) inhibits T cell activation. Antibodies to these receptors are currently frequently leveraged in combination with other cancer immunotherapy modalities to enhance anti-tumor T cell activity. Figure 1 retrieved without modifications from Morante et al. 2022.1Deed - Attribution 4.0 International - Creative Commons
To date, several immune checkpoint inhibitors in the form of monoclonal antibody drugs have been approved for various indications, including but not limited to metastatic melanoma, renal cell carcinoma, and non-small lung cancer.2
Checkpoint Inhibitor Target
Antibody Drug Name
PD-1
Nivolumab, Pembrolizumab, and Cemiplimab
PDL-1
Atezolimumab, Durvalumab and Avelumab
CTL-4
Ipilimumab
Information retrieved from Shiravand et al. 2022.2
Beyond PD-1, PDL1, and CTL-4, other immune checkpoints with therapeutic potential have been identified, including Lymphocyte activation gene-3 (LAG-3), T-cell immunoglobulin and mucin domain-containing protein 3 (TIM-3), V-domain Ig suppressor of T cell activation (VISTA), and more.3 Monoclonal antibodies targeting some of these more recently characterized immune checkpoints are undergoing clinical evaluation as monotherapies and in combination with other inhibitors or chemotherapy.3
Immune Checkpoint
Expression
Ligands-Immunosuppression
V-domain Ig suppressor of T cell activation (VISTA)
CD4+ and CD8+ T, Tregs, Dendritic and NK cells, monocytes, macrophages and granulocytes
V-set and Ig domain-containing 3 (VSIG3) and P-selectin glycoprotein ligand 1 (PSGL-1)4
B7 homolog 3 (B7-H3)
APCs, NK, B and T, tumor cells, ECs, and fibroblasts6
*Interleukin 20 receptor subunit alpha, and phospholipase A2 receptor 15
Lymphocyte activation gene-3 (LAG-3)
T (eff and reg); NK, B, and Dendritic, cells7,8
major histocompatibility complex class II (MHC II), LSECTin, fibrinogen-like protein 1 (FGL1), and Galectin-37,9
T-cell immunoglobulin and mucin domain-containing protein 3 (TIM-3)
CD8+ and CD4+ T helper 1, Treg, NK and Dendritic cells, monocytes and macrophages
Galectin-9, Ceacam1 and HMGB1
T-cell immunoreceptor with Ig and ITIM domains (TIGIT)
T (eff, mem, reg), T fh and NK cells
CD155 and CD112
Information retrieved from Chretien et al. 2019 and Yuan et al. 2021.3,4 Abbreviations: antigen-presenting cells-APCs, eff-effector, endothelial cells-ECs, fh-follicular helper, mem-memory, reg-regulatory. *Potential co-inhibitory receptors identified to date.5
Antibody-drugs targeting LAG-3
Among the novel immune checkpoints, accumulating evidence supports the blockade of the lymphocyte-activation gene-3 (LAG-3) receptor for cancer immunotherapy. Significantly, the phase 2/3 clinical study RELATIVITY-047 demonstrated that combining Relatlimab and Nivolumab significantly benefited the progression-free survival of patients with untreated advanced melanoma.7 These findings drove the FDA approval of Relatlimab, a humanized IgG4 monoclonal antibody, representing the third type of antibody drug targeting an immune checkpoint available for clinical use.7
Intense interest in LAG-3 as a valuable immunotherapy target in solid tumors continues to drive antibody discovery and development efforts. To date, several monoclonal antibodies (e.g., Favezelimab, Fianlimab, and Leramilimab) targeting LAG-3 and bispecific antibodies against LAG-3 and PD-1 (e.g., Tebotelimab and RO7247669) or LAG-3 and CTLA-4 (e.g., Pavunalimab) are undergoing clinical evaluation.7
Basis of LAG-3-mediated tumor immunosuppression
LAG-3 is broadly expressed in immune cells, including CD4+ and CD8+ T cells. Similar to the immune checkpoint PD-1, LAG-3 is not present on the surface of naïve T cells, rather, its expression is dependent on T cell activation.8 Within the tumor microenvironment, LAG-3 is found overexpressed on infiltrating T-cells (e.g., CD8+ T and Tregs) in several solid tumors, including renal cell carcinoma (RCC), hepatocellular carcinoma (HCC), melanoma, and more.7
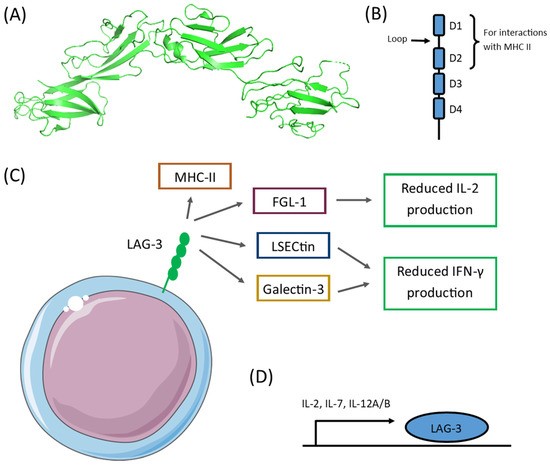
LAG-3 structure and ligands. A, B. LAG-3 is a Type-I transmembrane protein having four extracellular Ig-like domains with a 20% homology to the T cell CD4 receptor. Among these Ig-domains, D1-D2 are critical for interactions with MHCII. C. LAG-3 binds to other molecules relevant for immunosuppression, leading to reduced IL-2 and IFN-g production. D. Several cytokines, such as L-2, IL-7, and IL-12, have been shown to induce LAG-3 expression.7,8 Retrieved from Sauer et al. 2022 without modifications.9https://creativecommons.org/licenses/by/4.0/
LAG-3 overexpression in tumors creates a permissive environment that facilitates tumor survival, growth, and metastasis.7 To date, several LAG-3 ligands have been identified that help mediate its immunosuppressive effects. Moreover, LAG-3-dependent immune inhibition has been shown to occur through various processes, including inducing the exhaustion of effector T cells, promoting Treg differentiation, reducing NKT cell activity, inducing iNKT cell exhaustion, and inhibiting the activation and expansion of plasmacytoid dendritic cells.7
How do anti-LAG-3 antibodies work for immunotherapy?
Most anti-LAG3 candidates developed to date are blocking antibodies that inhibit binding to MHCII molecules.9 Blockade of LAG3 frees MHCII molecules to bind T-cell receptors and CD4, restoring downstream signaling to promote CD4+ T-cell proliferation and activation.7-9 Despite the benefits of anti-LAG-3 antibodies, approaches combining LAG-3 and PD-1 blockade are more therapeutically effective by also increasing CD8+ T-cell proliferation and activity.9
A new LAG-3-dependent mechanism of immune suppression
Various molecular mechanisms have been proposed for LAG-3's effects on T-cell activity. Among them, inhibitory signals transduced through the LAG-3 cytoplasmic domain have been shown to play an intrinsic role in T-cell suppression. Now, a study led by Tadashi Yokosuka at the Department of Immunology, Tokyo Medical University, has revealed a new extrinsic mechanism underscoring LAG-3-dependent immunosuppression.10
By leveraging various molecular imaging analysis strategies, the Yokosuka team has monitored and dissected molecular events at T-cell immunological synapses. First, in vitro studies in which T-cells were exposed to MHCII molecules anchored within a synthetic lipid bilayer enabled the team to visualize LAG-3 dynamics and test elements critical for its interaction with the MHCII complex. Using internal reflection fluorescence (TIRF) microscopy, they could visualize the clustering of LAG-3 within areas of contacts established between T-cell receptors and MHCII molecules at immunological synapses, co-localizing within “supramolecular activation complexes.” Within these complexes, the binding of LAG-3 with MHCII molecules was dependent on its first extracellular Ig domain (D1) and independent of the identity of the MHCII-bound peptide.
It has been previously established that within “supramolecular activation complexes,” TCR-bound MHCII molecules are conventionally internalized as part of the TCR-complex activity and homeostasis.11 Significantly, Yokosuka’s team found that LAG-3 could amplify the trans-endocytosis of peptide-MCHII complexes beyond those specifically bound to TCRs. This is because, upon T-cell and APC interaction, LAG-3-mediated MHCII internalization occurs independently from its haplotype and peptide specificity. Ultimately, the team found that this LAG-3-mediated enhancement in MHCII trans-endocytosis reduced overall MHCII expression on APCs.
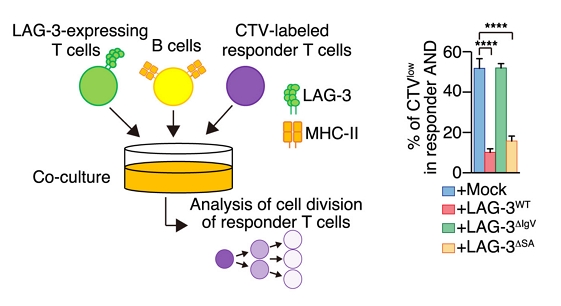
In vitro experimental design to evaluate LAG-3-mediated T cell immunosuppression. (Left) LAG-3 AND T cells and antigen-presenting B cells (pre-pulsed with the corresponding peptide MCC88–103 peptide) were co-cultured with CellTrace Violet (CTV)-labeled AND T cells as responder cells. (Right) The proliferation of CTV-Labeled AND T cells was significantly reduced only when co-cultured with T cells expressing wild-type LAG-3 or LAG-3ΔSA (i.e., missing a suppressive sequence in its cytoplasmic domain). In contrast, co-culture with unmodified (Mock) T cells or those expressing a mutant form of LAG-3 that cannot bind to MHCII molecules (LAG3ΔIgV) did not diminish the proliferation of CTV-Labeled AND T cells. Retrieved with modifications from Wakamatsu et al. 2024, only Figure 4’s panel A and a portion of panel B are shown. Deed - Attribution 4.0 International - Creative Commons
This finding raised the question of whether LAG-3-mediated MHCII trans-endocytosis could induce substantial immune suppression. Through in vitro experiments with CD4+ T cells under co-culture conditions with peptide-loaded APCs and corresponding antigen-specific LAG-3-expressing T cells, Yokosuka and colleagues demonstrated that LAG-3 can indeed reduce T cell proliferation through its indirect effects on APCs. Significantly, the team found that CD8+ T-cells expressing LAG-3 can also mediate MHCII trans-endocytosis and reduce antigen presentation, consequently reducing CD4+ T-cell activation.
Conclusion
Overall, LAG-3 is broadly expressed in immune cells, and its inhibitory mechanisms are not yet fully elucidated. As monoclonal antibody drugs targeting LAG-3 advance through clinical studies, a deeper understanding of the mechanisms underscoring their intrinsic and extrinsic effects is critical, particularly within the context of combined therapies with other immune checkpoint inhibitors.
References
1. Morante M, et al. 2022. Immune Checkpoint Inhibitors and RAS–ERK Pathway-Targeted Drugs as Combined Therapy for the Treatment of Melanoma. Biomolecules. https://doi.org/10.3390/biom12111562
3. Chrétien S, et al. 2019. Beyond PD-1/PD-L1 Inhibition: What the Future Holds for Breast Cancer Immunotherapy. Cancers. https://doi.org/10.3390/cancers11050628
4. Yuan L, et al. 2021. VISTA: A Mediator of Quiescence and a Promising Target in Cancer Immunotherapy. Trends Immunol. https://doi.org/10.1016/j.it.2020.12.008
7. Ibrahim R, et al. 2023. LAG-3 Inhibitors: Novel Immune Checkpoint Inhibitors Changing the Landscape of Immunotherapy. Biomedicines. https://doi.org/10.3390/biomedicines11071878
9. Sauer N, et al. 2022. LAG-3 as a Potent Target for Novel Anticancer Therapies of a Wide Range of Tumors. International Journal of Molecular Sciences. https://doi.org/10.3390/ijms23179958
10. Wakamatsu E. et al. 2024. Indirect suppression of CD4 T cell activation through
LAG-3-mediated trans-endocytosis of MHC class II. Cell Reports. https://doi.org/10.1016/j.celrep.2024.114655
 Most Popular Products
Most Popular Products
 CRISPR Gene Editing
CRISPR Gene Editing
 Antibodies
Antibodies
 Protein Electrophoresis and Blotting
Protein Electrophoresis and Blotting
 Molecular Biology
Molecular Biology
 Stable Cell Lines
Stable Cell Lines
 Cell Therapy
Cell Therapy
 Diagnostics
Diagnostics
 Resources
Resources